Как хорошо на свете жить,
С друзьями вместе водку пить,
Всем им
рассказывать муру́,
И вешать на уши лапшу́ ))
|
* В генетической генеалогии звёздочкой {*} обозначается исходная гаплогруппа, от к-рой пошли дочерние гаплогруппы. Например,
от материнской C* произошли дочерние C1 и C2. C без звёздочки обозначает обобщение C1 и
C2, в свою очередь являющихся материнскими для происходящих от них гаплогрупп, обозначаемых чередованием латинских
букв и арабских цифр )), например C1a2.
** Время обозначается в тыс. лет до н.э., например, 88 т. значит 88 тысяч лет до нашей эры. |
| A– койсаны (гаплогруппу A имел ещё хромосомный Адам в 250 т.) | |||||||||||||||||||||
| BT | {130 т. или 70-80 т.} – найдена у народности хуэй в Китае | ||||||||||||||||||||
| B | {88 т.} - пигмеи | ||||||||||||||||||||
| CT | {88 т.} – существовала до «исхода из Африки», видимо, на её севере | ||||||||||||||||||||
| DE | {68 т.} – носители парагруппы DE* очень редки, есть в Африке и Азии, следовательно она возникла в Африке и распалась в Азии | ||||||||||||||||||||
| D | {50 т.} – азиатская ветвь: айны, андаманцы, много у японцев и в Тибете | ||||||||||||||||||||
| E | {50 т.} – африканская ветвь: негры, есть субклады E1, E2, …; E1b1* {41-49 т.} найдена в Эфиопии; очень многочисленны: | ||||||||||||||||||||
| E1b1a | {42 т.} - банту | ||||||||||||||||||||
| E1b1b | {42 т.} - нилоты | ||||||||||||||||||||
| CF | {66 т.} – носители CF* не обнаружены, видимо, она тоже возникла в Африке и, вытеснив DE*, вслед за ней перебралась в Азию | ||||||||||||||||||||
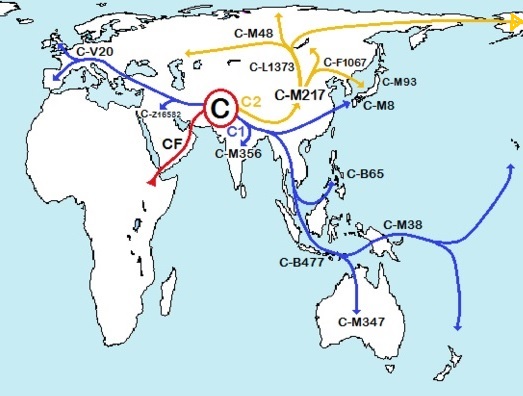
| C | {60 т.} – австралоиды*, часта на востоке Азии и у австралийских аборигенов | ||||||||||||||||||||
| F | {55 т.} – редкая линия гаплогруппы F, являющаяся внешней для остальной её части, найдена во Вьетнаме | ||||||||||||||||||||
| G {48 т.} - кавкасионы; G1 - вост.Кавказ, Ближний Восток, Ср.Азия; G2 - зап.Кавказ, юг Европы | |||||||||||||||||||||
| HIJK | {?} – HIJK* не найдена среди живых и мёртвых | ||||||||||||||||||||
| H | {48.5 т.} – веддоиды, цыгане | ||||||||||||||||||||
| IJK | {48.5 т.} - IJK* не найдена ни среди мёртвых, ни среди живых )) | ||||||||||||||||||||
| IJ | {43 т.} – кроманьонцы, IJ* не найдена среди живых и мёртвых | ||||||||||||||||||||
| I {20 - 25 т.} - нордиды; I1 - север Европы, I2 - Балканы | |||||||||||||||||||||
| J {31 т.} - арменоиды; J1 - Ближний Восток, север Африки, J2 - Анатолия, есть в Европе | |||||||||||||||||||||
| K | {47 т.} – K* найдена только у одной из таримских мумий | ||||||||||||||||||||
| K1 | =LT {41-49 т.} – прото-шумеры (в Джирофте) | ||||||||||||||||||||
| L {25 - 30 т.} – население Хараппы, дравиды | |||||||||||||||||||||
| T {30 т.} – шумеры, эламиты, финикийцы | |||||||||||||||||||||
| K2 | {45 т.} – K2* распространена у австралийских аборигенов (обозначается как K2-M526, её у них 29% **) | ||||||||||||||||||||
| K2a | {44 т.} – предок монголоидов, K2a* найдена у усть-ишимского человека и в Пештера-ку-Оасе | ||||||||||||||||||||
| K2a1 | =NO {43 т.} – жёлтая раса, монголоиды | ||||||||||||||||||||
| N {34 т.} – северные монголоиды, балты, угро-финны*** (у венгров почти не осталось) | |||||||||||||||||||||
| O {34 т.} – южные монголоиды – аустрики, китайцы*4, монголы, корейцы, японцы | |||||||||||||||||||||
| K2b*5 | {44т.} – исходная K2b*, так же как K2b1*, не найдены | ||||||||||||||||||||
| K2b1 | – субклады (в т.ч. M и S) – аэта (Филиппины) 60%, Соломоновы о-ва 72%, Папуа 83% | ||||||||||||||||||||
| M=K2b1b {32 -47 т.} – прибрежные папуасы | |||||||||||||||||||||
| S = K2b1a { 28 - 41 т.} – горные папуасы, меланезийцы, есть в Австралии | |||||||||||||||||||||
| K2b2 | = P {34 т.} – P* редка, найдена в южной, юго-восточной, восточной Азии и в Океании | ||||||||||||||||||||
| K2b2a | =P1 {31 т.} – аэта 28%, австралоиды 0, Папуа 0.89% | ||||||||||||||||||||
| K2b2a1 | = Q {15-20 т.} – кеты*6, индейцы, гунны | ||||||||||||||||||||
| K2b2a2 | = R {24-34 т.}*7-древние средиземноморцы; R*-10.3% у буришек и 6.8% у калашей | ||||||||||||||||||||
| R1 {25-30 т.}*8 - до 5 т. - в Малой Азии и Леванте | |||||||||||||||||||||
| R1a {20 т.}*9 – в восточной Европе | |||||||||||||||||||||
| R1b {20 т.}*10 – в западной Европе и сев.Африке | |||||||||||||||||||||
| R2 {30-35 т.} - до 5 т. - в сев. Месопотамии и Иране, сейчас в Индии и её западных окрестностях )), особенно у дравидов | |||||||||||||||||||||
| K2b2b | = P2– очень редка, найдена на о-ве Лусон у аэта | ||||||||||||||||||||
| K2c, K2d, K2e – очень редки, найдены в Индии и Индонезии | |||||||||||||||||||||
| - Барриноидный тип (включая тасманийцев?), близки к меланезийцам, грацильные темнокожие курчавые низкорослые, жили в дождевых лесах севера Австралии, где укрылись от последующих пришельцев и измельчали. Папуасские гаплогруппы M и S есть в Австралии, 1-я редка, но распространена по всему материку, 2-я наиболее распространена на юге. | Последние исследования однако указывают на то, что эта группа представляет собою локальную модификацию австралоидного типа [3]. | ||
| - Мюррейский тип, имеет айнский компонент (?), кожа и глаза светлее, очень волосатые, крупная голова и зубы, средний рост, жили на юге (гаплогруппа К2-М526 ?). | Тасманийцев сближают и с мюррейским типом. У тасманийцев был очень маленький череп - меньше только у андаманцев, но зубы очень большие - почти как у австралийских аборигенов - как это приятно звучит для уха расиста )). | ||
| - Карпентарийский тип, очень чёрные, узенькие и очень высокие, волосатость низкая, жили на севере и в центре (преобладает гаплогруппа С1 ). | |||
| Соответственно, эти переселения произошли в ~ 40, 20, 15 т. | |||
|
Вопрос о происхождении тасманийцев м.б. решён исследованием палеоДНК или особенностей генотипа европейско-тасманийских метисов,
но таких работ пока нет [4]. Создаётся впечатление о близости тасманийцев
с андаманцами и айнами, следовательно, у них д.б. гаплогруппа D. | |||
|
В целом, надо полагать, в Австралию 1-ми прибыли айны гаплогруппы D, положившими начало мюррейскому типу на юго-востоке
Австралии и * А вот сами папуасы почему-то не измельчали )) | |||
|
| |
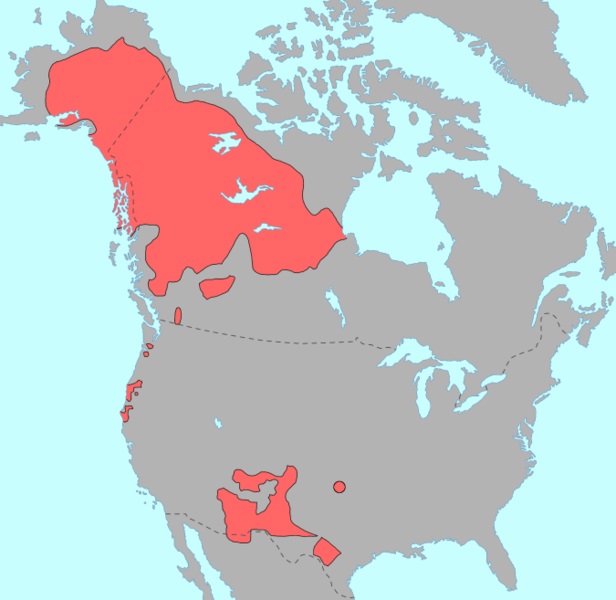
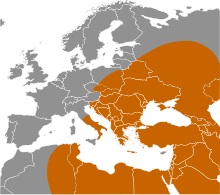
| Распространение по белу свету гаплогруппы C )) | Распространение языков на-дене в Северной Америке )) |
| Рис.2 | |
| ЗАМЕЧАНИЕ. Изначально южные монголоиды гапллогруппы O говорили на аустрическом языке, к-рый у многих южных и почти всех северных монголоидов был заменён на языки сино-кавказской или ностратической макро-семей. Но не сохранились ли реликтовые языки северных монголоидов? Возможно, таким языком является чукотский - у чукчей гаплогруппы N = 61%, а родственный чукотскому ительменский язык является заимствованным - у ительменов гаплогруппы C = 67%, они австралоиды. Тогда нивхи*, у к-рых гаплогруппы C = 71%, говорят на языке северных австралоидов. | ||
| * С их соседями удегейцами всё ясно - у них язык тунгусо-маньчжурской семьи. | ||
|
И ещё. Язык кетов принадлежит к сино-кавказской макро-семье, у кетов основной является гаплогруппа Q, также как у
американских индейцев. Следовательно, язык кетов заимствован, а америндские языки индейцев изначальны для гаплогруппы
Q и родственны ностратическим, на к-рых говорят носители гаплогруппы R. Вот к каким лингвистическим выводам приводит исследование гаплогрупп! А подробнее об этом - см.здесь | ||
|
* Языки нивхов и юкагиров в отдалённом родстве, они говорят на изначальном языке гаплогруппы C2; удэгейцы перешли на тунгусский язык, коряки - на чукотский ** Эта гаплогруппа была у Чингис-хана *** Эта гаплогруппа - у маньчжурской династии Цин в Китае |
|
* Это справедливо как для одиночного хищника - например, тигра, так и прайда львов или стаи волков.
** Именно поэтому передающиеся по женской линии митохондриальные гаплогруппы сейчас мало-информативны - они крайне перемешались ещё в древности. |
| - молодёжь самнитов положила начало племенам бруттиев и луканцев в южной Италии; | |
| - молодёжь германцев, теснимых римскими легионами, под именем маркоманнов положила начало г-ву Маробода в Чехии; | |
| - молодёжь Афин под предводительством сыновей последнего афинского царя Кодра основала греческие колонии в Милете и Эфесе, перебив местное мужское население, о чём см.здесь |
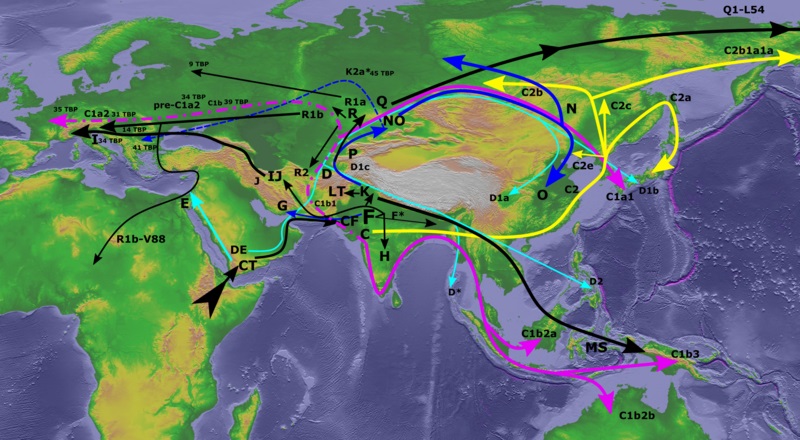 Рис.3 |
* Судя по гаплогруппам, первобытные охотники и собиратели предпочитали не смешиваться, а истреблять друг друга. Видимо, уже тогдашний мир был перенаселён человеком, ресурсов не хватало. Правда, наличие сексуальных контактов кроманьонцев и неандертальцев не подлежит сомнению - видимо, и тогдашних людей тянуло на клубничку )).Представители CF тоже перебрались на Ближний Восток. Здесь они энергично теснили D, истребляя задержавшихся - следов D в Индии не обнаружено. Остатки D укрылись на Андаманских о-вах, холодном севере Восточной Азии - айны*, малодоступном Тибете и, возможно, в Тасмании (см.здесь).
** При этом они так спешили, что не успели сойтись с ближне-восточными неандертальцами )) - в Африке нет неандертальских генов.
*** Это айны, они имеют гаплогруппу D и представляют, следовательно, особую древнюю расу.
*4 Следовательно, показанный на карте голубой стрелой северный маршрут расселения гаплогруппы D неверен. Она расселялась южнее, через Индию, Индокитай и Китай, так же как расселялась по жёлтой стреле гаплогруппа C.
* Айны доставили D в Японию, куда преследовавшие их австралоиды C проникнуть не смогли - видимо, поднялся уровень океана.Австралоиды C заселили Малую Азию, Иран, Индию, Индокитай, Китай, позже через Индонезию достигли Австралии* - по большей части посуху, ибо уровень мирового океана тогда был намного ниже современного. Так гаплогруппа CF разделилась на C австролоидов и оставшуюся пока в Передней Азии F. Вскоре и она начала экспансию, тесня и изгоняя своих родственников C. Основной поток F шёл на восток, по тропическому поясу, но часть их подавалась на север, на новые неизведанные земли. Там стали возникать новые гаплогруппы.
| * Распространяясь, носители гаплогруппы F отрезали от основного массива C её западную ветвь C1a, находившуюся в Малой Азии. |
* Строго говоря, в кроманьонцах состояли как носители
IJ, так и C1a. Последняя найдена у представителей Ориньякской к-ры 33-24 т. - в пещере Гримальди на западе
ареала, а на Костёнковских стоянках Воронежской области, на востоке ареала этой к-ры, найдены C1b и даже CT.
Но, безусловно, носители IJ преобладали.  |
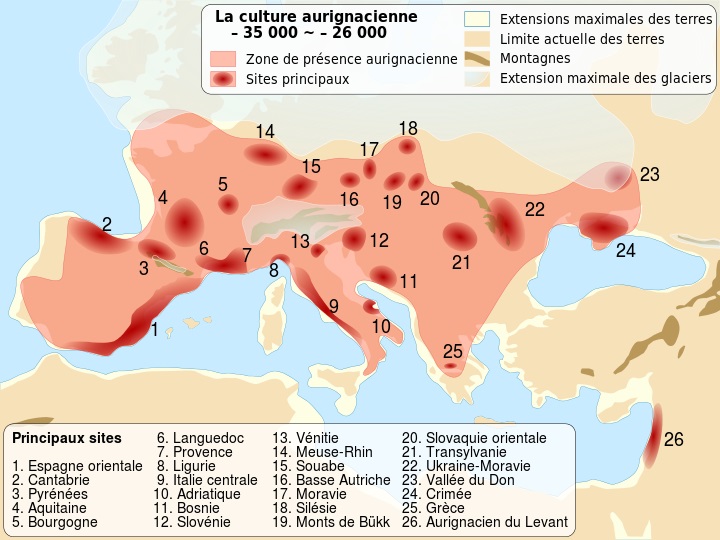 Ориньякская к-ра 33 - 24 т. - имеются и другие её датировки |
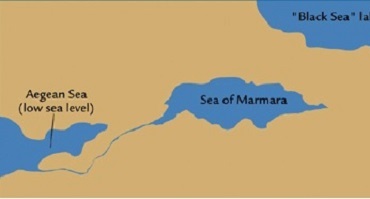
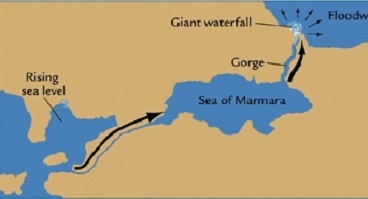
| ** Лишь ок. 5.6 т. из-за землетрясения Босфорский перешеек
был прорван***, в результате катастрофического наводнения Чёрное море заполнилось солёной водой, что показано на
рис.5. *** Это событие, видимо, нанесло страшный удар процветающей в Малой Азии цивилизации средиземноморцев гаплогруппы R1a - города Чатал-Гуюк, Хаджилар и др., что и позволило | |
| кавкасионам гаплогруппы G начать свои завоевания на Ближнем Востоке, положившим начало Халафской к-ре 5.6 - 4.5 т., приведшим к вытеснению средиземноморцев в Европу и афразийцев R1b в сев. Африку, распространению по миру ностратических языков. | |
| Рис.4 | |
 |  |
 |  |
| Прорыв вод мирового океана в Чёрное море | Человек из Костёнки 14* |
Айн гаплогруппы D | |
| * Он жил ок. 37 т., похож на | |||
| Рис.5 | |||
| Как кроманьонцы заселяли Европу | ||||
 |  |
 |  |  |
| к 37.5 т. | к 35.0 т. | к 32.5 т. | к 30.0 т. | Юноша-кроманьонец |
| из Сунгири 2 Владимирской обл. жил ок. 34 т. | ||||
| Рис.6 | ||||
| * Койсаны в пустынях юга Африки подобно монголоидам пожелтели, но остались чернокожими - ведь с неандертальцами они не сталкивались, избавиться от "чёрных" генов не так-то легко )) |
| * Соответственно, G двигалась на север, IJ на запад, K на восток, а H хоть и не сражалась, но оставалась победителем на поле сражения )) |
| - 1-й - через Среднюю Азию, обходя Тибет с запада, как это показано на карте; | |
| - 2-й - через Китай, обходя Тибет с востока. |
| 2 варианта движения на север гаплогруппы K2 | |||
| 1-й маршрут - через Среднюю Азию | 2-й маршрут - через Китай | ||
| В соответствии с картой, часть гаплогруппы K из Индии вошла в Среднюю Азию, где часть её, именуемая K1 =
LT - будущие прото-шумеры остались, а другая K2 движение на север, в Великую степь, 2-мя волнами: 1-й шла
K2b, её подпирала Ka.
K2b разделилась на K2b1 = MS и каким-то образом достигла Новой Гвинеи, а K2b2a = P1 в Великой степи стала расходиться: на запад гаплогруппа R, на восток - гаплогруппа Q, причём последняя начала желтеть. Обеих всё больше разгоняла вклинившаяся меж ними гаплогруппа K2a = NO, так же пожелтевшая в Великой степи. Исходная же гаплогруппа K* продолжала идти на восток, через Индию и Индокитай, и в конце концов поселилась в Австралии. Прим. Непонятно только, каким образом гаплогруппа P2, сестринская для P1, добралась до острова Лусон, как сама P1 оказалась у аэта, как MS достигла Новой Гвинеи. |
Оставив K1 = LT в Средней Азии, гаплогруппа K достигла Индо-Китая, где на какое-то время поселилась
её исходная гаплогруппа K*. Выделившаяся из неё передовая K2 начала дробиться:
субклада K2b разделилась: на K2b1 = MS, к-рая направилась в Новую Гвинею, и на K2b2 = P, к-рая
тоже поделилась: K2b2b = P2 осталась на месте, а K2b2a = P1 = RQ
подалась через Китай на север, подпираемая следующей за ней субкладой K2a, от к-рой исходит исходная для
монголоидов гаплогруппа K2a1 = NO.
В Великой степи P1 распалась на субклады R и Q: 1-я пошла на запад*, 2-я на восток, желтея в восточной полупустынной части Великой степи. Разрыв между ними расширялся вклинившейся субкладой NO, гнавшей R в Европу, а Q - в Сибирь и Америку. Сама NO, тоже пожелтевшая, распространилась по всей Великой степи - её представители найдены на усть-ишимской стоянки Омской области ок. 45 т. и в карпатской «Пе́штера-ку-Оа́се» Румынии ок. 40 т. Сама она распалась на северо-западную N и южную O. Последняя во время ледникового периода хлынула на юг, ассимилируя местное чернокожее население гаплогрупп C, K*, MS, P2. * У мальчика с палеолитической стоянки Мальта Иркутской области найдена исходная гаплогруппа R* - это наиболее восточный пункт распространения гаплогруппы R и подтверждает, что на север P1 шла через Китай Вот так сформировалась современная жёлтая раса. | ||
| Неувязки 1-го маршрута дают основание отдать предпочтение 2-му | |||
  |
 |
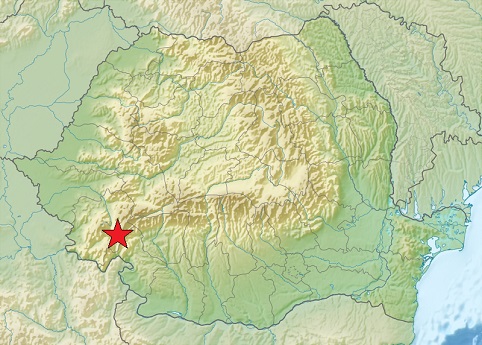
| Вот так выглядел предок монголоидов, живший 40 тыс. лет тому назад в карпатской пещере «Пе́штера-ку-Оа́се» Румынии. Он обладал гаплогруппой K2a*, исходной для монголоидной гаплогруппы K2a1 = NO. В нём ещё видны негроидные черты. На карте отмечено место его проживания | А это его ближайший родственник, проживавший на усть-ишимской стоянке омской области. Он обладал той же гаплогруппой и жил 45 тыс. лет тому назад. Негроидности в нём куда больше |
| Рис.7 | |